What Is The Role Of Thymic Stromal Lymphopoietin In Respiratory Disorders?
The thymic stromal lymphopoietin in respiratory disorders is about the protein (TSLP) produced by cells in the lungs and other tissues. It triggers inflammation and promotes immune responses.
Author:Suleman ShahReviewer:Han JuSep 12, 20241.7K Shares431.8K Views

The aim of this review is the assessment of thymic stromal lymphopoietin in respiratory disorders, with a focus on sinonasal diseases.
The epithelial cells of airways, skin and gut represent the first barrier of the human body against foreign antigens.
Several recent studies demonstrated the pivotal role of epithelial derived cytokines in driving the immune response.
Here we review thymic stromal lymphopoietin (TSLP) structure, its interaction with the TSLP receptor (TSLP R), its tissue expression and signaling mechanism.
Moreover, its actions on priming dendritic cells and polarizing T cells have also been reviewed, with a focus on the role TSLP plays in chronic inflammatory disorders of the upper respiratory tract.
Discussion
The role of thymic stromal lymphopoietin (TSLP) consists mainly in polarizing dendritic cells to induce the differentiation of naïve T cells into inflammatory T helper 2 (Th2) cells by up-regulating co-stimulatory molecules and OX40 ligand.
Moreover, TSLP is involved in innate immunity response by promoting cytokine secretions from mast cells, eosinophils and macrophages. TSLP and other epithelial-derived cytokines (IL25 and IL33) constitute a wide and complex network for the regulation of immunity, inflammation, and immune regulation.
All these data indicate that epithelial cells may have a critical role in the initiation of Th2 immune responses at mucosal sites through TSLP, IL25 (interleukin-25) and IL33 (interleukin-33) production, acting not only like a physical but also like a functional barrier.
TSLP Cytokine
The epithelial cell-derived thymic stromal lymphopoietin (TSLP) was first identified in 1994 in supernatants from mouse thymic stromal cell lines Z210R.1.
Further molecular characterization in humans showed human TSLP had only 43% amino acids homology with mouse TSLP, with similar biological functions and tissue localization.
The human TSLP gene is located on chromosome 5q22 not far from the cluster gene encoding the Th2 cytokines.
TSLP is a 140-amino acid IL-7-like 4-helix bundle cytokine mainly expressed by barrier ECs in the thymus, lung, skin, bowel and tonsils and has been recently identified as a key factor in Th2-inflammatory response.
TSLP is constitutively expressed in intestinal and thymic epithelial cells.
TSLP is involved in tolerance to commensal flora in the bowel and in differentiation of T regulatory cells in the thymus, by modulating the dendritic cells activity.
TSLP expression may be induced by different exogenous/endogenous stimuli as well as by:
- pathogens
- traumas
- infections
- allergens
- Toll-like receptor ligands
- pro-inflammatory
- Th2 cytokines
Thymic Stromal Lymphopoietin Receptor
Thymic stromal lymphopoietin receptor (TSLP R) complex is made up of a TSLP-binding chain and the interleukin 7 receptor α (IL7Rα) chain.
Human and mouse TSLP R share about 40% of the protein sequence.
The human TSLP R gene is located on chromosome Xp22.3 and Yp11.3.
TSLP R is expressed on several immune cells:
- early B and T cell progenitors
- peripheral CD4+ T cells
- natural killer T cells (NKT)
- monocytes
- mast cells
- dendritic cells
It is also expressed on cells found in the:
- heart
- skeletal muscle
- kidney
- liver
In humans, thymic stromal lymphopoietin receptor (TSLP) activates the signal transducer and activator of transcription (STAT)-5 and (STAT)-3 by interacting with the heterodimeric complex TSLP R/IL7Rα (interleukin-7 receptor alpha).
It has been shown that TSLP and IL7 receptors use different signal mechanisms to activate STAT-5. IL7 activates JAK1 (Janus kinase 1) and JAK3 (Janus kinase 3), while TSLP was believed not to activate any JAK kinases.
In contrast to this view, two recent studies showed activation of JAK1 and JAK2 after TSLP signaling in both human dendritic cells and human CD4+ T cells, aka CD4 cells.
JAK1 and JAK 2 are synergically bound from the IL7Rα and the TSLP R subunits, and, in addition to STAT 5, TSLP stimulation can activate:
| STAT1 | STAT6 |
| STAT3 | JAK1 |
| STAT4 | JAK2 |
| STAT5 | -- |
Presumably, other signaling pathways are activated by TSLP, such as:
- PI3-K/Akt (phosphoinositide 3-kinases)
- MAPK (mitogen-activated protein kinases)
- Src family Kinase pathways
TSLP And Immune Response
The role of thymic stromal lymphopoietin (TSLP) consists mainly in polarizing dendritic cells to induce the differentiation of naïve T cells into inflammatory Th2 cells by up-regulating:
- co-stimulatory molecules CD40 (cluster of differentiation 40) and CD80 (cluster of differentiation 80)
- MHC (major histocompatibility complex) II molecules
- OX40 ligand (OX40L)
Moreover, TSLP is involved in innate immunity response by promoting cytokines secretions from:
- mast cells
- eosinophils
- macrophages
Dendritic Cells
Dendritic cells are professional antigen presenting cells (APCs) which may be considered a bridge between innate and adaptive immunity.
The maturation of human myeloid dendritic cells can be enhanced by TSLP; and high level expression of TSLP R on these cells allows a rapid response to TSLP, up-regulating:
- MHC class II
- CD40, CD80, and CD86
- OX40 ligand (OX40L)
- TARC (thymus and activation-regulated chemokine)
- MDC (macrophage derived chemokine) molecules
TSLP-treated dendritic cells up-regulate OX40L, do not produce IL12, and drive naïve CD4+ T cells to differentiate into inflammatory Th2 cells that produce Th2 cytokine like IL4, IL5, IL13 and large amount of TNF alpha, but not IL10.
Dendritic Cells And OX40/OX40L
OX40 ligand (OX40L), originally described as a glycoprotein (termed gp34), and its receptor (OX40), belongs to the superfamily of TNF (tumor necrosis factor)/TNF receptor.
OX40L is preferentially expressed on antigen presenting cells like:
- myeloid
- plasmacytoid dendritic cells (pDCs)
- B cells
It can also be expressed on:
- vascular endothelial cells
- mast cells
- NK (natural killer) cells
OX40 is mainly expressed by activated CD4 Tcells, 24 to 48 hours after engagement of T cell receptor (TCR), and not constitutively expressed on naïve T cells.
According to some observations, OX40/OX40L interaction is involved in Th2 inflammatory diseases, particularly allergic inflammation, and its role is crucial for T cell activation and polarization towards a Th2 inflammatory pattern.
High levels of OX40L are expressed by TSLP-activated dendritic cells, which play an important role in driving Th2 cells expansion by interacting with OX40+ CD4 T cells.
TSLP And Adaptive Immunity
Thymic stromal lymphopoietin (TSLP) acts on CD4+ T cells, not only indirectly through dendritic cells and OX40L, but also in a direct way.
In vitro studies demonstrated that T cell receptor (TCR)-engagement and consequent activation of T cells induce TSLP R expression and proliferation of pre-activated CD4+ T lymphocytes leading to their responsiveness to TSLP stimulation.
T regulatory cells (CD4+/CD25+/Foxp3+) are a small population of T cells with a suppressive and regulatory function, which are thought to be important in the maintenance of tolerance towards self antigens.
Their differentiation is under the control of the transcription factor forkhead box protein 3 (FOXP3).
In thymic medulla, approximately 50% of CD4+/CD8-/CD25-thymocytes bearing TCR with high affinity for self peptide-MHC (major histocompatibility complex) complexes are induced by Hassall’s corpuscles (HCs) express thymic stromal lymphopoietin (TSLP) to differentiate into Treg cells in a dendritic cell-dependent manner.
TSLP action is also explicated on activation and differentiation of CD8+ T cells into pro-allergic cytotoxic T cells that produce large amounts of IFN-γ (interferon gamma).
Similar to CD4+ T cells, TSLP can induce TSLP R expression on TCR-activated CD8+ T cells increasing their survival through Bcl-2 (B-cell lymphoma 2) anti-apoptotic protein.
The role of TSLP on B cells development is still unclear: in vitro studies suggested that TSLP can contribute to B lymphopoiesis, being active on pro-B cells derived from fetal liver.
In vivo results are contradictory about the role of TSLP in B cells development, as it has been demonstrated in a study published in 2004 by the journal Molecular and Cellular Biology, with Nick Carpino as lead author.
Based on that study, B cell and T cell developments are normal in TSLPR (-/-) mice.
This discrepancy can be the result of different levels of circulating TSLP in transgenic compared to wild mice.
TSLP And Innate Immunity
Thymic stromal lymphopoietin (TSLP) can also influence cells of innate immunity like:
- mast cells
- basophils
- eosinophils
- natural killer T cells (NKT cells)
NKT cells, like the other T cells, develop from thymocytes progenitors and after TCR activation can rapidly produce IL-4 and IFN-γ (interferon‐gamma).
High percentages of NKT cells have been found in bronchoalveolar lavage fluid (BALF) in asthmatic patients with airway hyperresponsiveness (AHR). It has been observed that TSLP, in these patients, may induce NKT cells to produce IL-13, which, in turn, is able to increase airway inflammation and reactivity.
Mast cells, basophils, and eosinophils are cells that commonly contribute to immediate hypersensitivity reactions in allergic responses.
Mast cells are one of the identified non-epithelial sources of TSLP.
Following immunoglobulin E (IgE) receptor activation, mast cells are able to produce TSLP and up-regulate the expression of TSLP R complex, leading to TSLP response in a pro-inflammatory environment (IL1, TNF alpha).
TSLP stimulation of mast cells do not induce their degranulation, but it promotes the release of several:
a. Th2 cytokines:
- interleukins (IL) - IL5, IL6, IL13
- tumor necrosis factor (TNF) alpha
- GM-CSF (granulocyte-macrophage colony-stimulating factor)
b. chemokines:
- CCL1 or chemokine (C-C motif) ligand 1
- CXCL8 or chemokine (C-C motif) ligand 8
The pivotal role of mast cells in the induction of TSLP production has been demonstrated in a study published in 2008 by the European Journal of Immunology, with Masanori Miyata as lead author.
Moreover, TSLP is able to activate common progenitors of basophils and eosinophils.
Basophils represent less than the 1% of circulating leukocytes, and they have been identified as a potential source of TSLP; so, contributing together with the other epithelial-derived cytokines to the Th2 allergic response.
Human eosinophils express the functional TSLP R complex and can be directly activated by TSLP in a dose-dependent and specific manner.
Many effects of TSLP on eosinophils have been reported:
- delayed apoptosis
- up-regulation of cell surface expression of adhesion molecule CD18 and intercellular adhesion molecule-1, with down-regulation of L-selectin
- enhanced adhesion to fibronectin
- augmented release of inflammatory cytokine IL-6 and chemokines CXCL8 (C-X-C Motif Chemokine Ligand 8), CXCL1, and CCL2
TSLP has been shown to regulate the above reported effects on eosinophils through the activation of extracellular signal-regulated protein kinase, p38 mitogen-activated protein kinase, and NF-kB (nuclear factor kappa B) signaling pathway, and not through STAT-5 and STAT-3, which are usually activated by TSLP stimulation of other effector cells.
Moreover, group 2 innate lymphoid cells (ILC2s) have been recently shown to play a critical role in the early phases of the development of allergic inflammation at multiple barrier surfaces.
The epithelial cell-derived cytokines IL-25, IL-33, and TSLP regulate the activation and effector functions of ILC2s.
TSLP And Epithelial Cell-Derived Cytokines Network
Several recent studies investigated the role of epithelial cells in licensing the functions of specific immune cell populations in the airway and gastrointestinal tract.
It has been demonstrated that epithelial cells are not only a physical barrier in the mucosal sites, but also play a key role in the initiation, regulation, and resolution of innate and adaptive immune responses.
TSLP and other epithelial-derived cytokines (IL25 and IL33) constitute a wide and complex network for the regulation of immunity, inflammation, and immune regulation.
This triad of cytokines share a common cellular source (epithelial cells) but they differ in termsof structure and cell population targets.
Tissue cytokines promote Th2 response in vivo and are important in modulating mucosal immune response.
TSLP, IL25, and IL33 are able, directly and/or indirectly, to initiate the inflammatory cascade, its progression, and regulation, and they seem to be involved in several diseases.
Interactions among tissue cytokines are regular and not entirely known.
IL25 and IL33 together are able to induce TSLP mRNA in epithelial cells.
These cytokines can up-regulate with a positive enhancing loop with their cognate receptors and IL33 is able to sensitize mast cells to TSLP.
Both TSLP and IL25 have many effects on antigen presenting cells (APCs) and CD4+ naïve T cells, the first one by up-regulating GATA3 (GATA-binding protein 3) expression and IL4 production, the second one by up-regulating IL4, IL5, and IL13 production and activating T CD4+ memory cells.
These three epithelial-derived cytokines have many effects on the innate cell populations of:
- eosinophils
- basophils
- mast cells
TSLP, in a pro-inflammatory environment (with or without IL33), promotes mast cells production of IL4, IL5, and IL13 and Th2-like chemokines.
IL33 can furthermore increase mast cells survival and sensitize them to TSLP stimulation. Basophils are induced to produce cytokines by IL33 and IL25 can increase eosinophils survival.
All these data indicate that epithelial cells may have a critical role in the initiation of Th2 immune responses at mucosal sites through TSLP, IL25, and IL33 production.
TSLP And Diseases
Many experimental data indicate that thymic stromal lymphopoietin (TSLP) plays a critical role in Th2-driven response.
Its increased production has been shown in target tissues of patients with Th2-related diseases like:
- atopic dermatitis
- asthma
- allergic rhinitis
Recently, TSLP has been found highly expressed in sinus mucosa of patients with chronic rhinosinusitis with nasal polyps (CRSwNP) and its role in cancer escape has been recently studied in patients with breast and pancreatic tumors.
TSLP And Atopic Dermatitis
Atopic dermatitis is a chronic inflammatory skin disorder of variable severity characterized by eczematous skin lesions associated with itching.
It has a strong genetic component and may affect:
- children (10% to 20%)
- adults (1% to 3%)
Atopic dermatitis is characterized by proliferation of epidermal keratinocytes and abnormal keratinization. Skin lesions contain immune infiltrates of Th2 cells and DCs.
High levels of total immunoglobulin E (IgE) and eosinophil numbers in the peripheral circulation are common in these patients.
High levels of thymic stromal lymphopoietin (TSLP) and inflammatory cytokines (e.g., IL-1b, TNF-alpha, IL-4, and IL-13) have been shown in keratinocytes from skin biopsy specimens of patients with atopic dermatitis, suggesting a feed-forward inflammatory cascade that synergize to induce TSLP expression by keratinocytes.
It likewise suggests that TSLP may play a role in the Th2 inflammatory aspect of this disease.
In murine models of atopic dermatitis, TSLPR (-/-) mice showed a significant decrease of skin infiltrating eosinophils and tissue T helper 2 (Th2) cytokines concentration, while intradermal injection of anti-TSLP antibodies were able to block the development of allergic skin inflammation.
TSLP has also been implicated in the phenomenon referred to as the atopic march, that refers to the chance of subjects with atopic dermatitis to develop allergic rhinitis and asthma in the future.
Several murine models of induced TSLP expression in keratinocytes result in allergic airway inflammation after intranasal challenge, suggesting that TSLP might be an important factor contributing to this progression from atopic dermatitis to allergic rhinitis and asthma.
TSLP And Asthma
Asthmais a disease characterized by chronic airway inflammation that is mediated by:
- T helper 2 (Th2) cells and their related cytokines
- immunoglobulin E (IgE) production
- innate immune cells recruitment
- mucus production
The inflammatory mediators released by activated mast cells in a IgE-dependent way are responsible for:
- an increased inflammatory cell infiltrate and mucus production
- bronchial smooth muscle contraction
- increased vascular permeability
Several recent studies suggest thymic stromal lymphopoietin (TSLP) may play a key role in the pathogenesis of allergic and non-allergic asthma, both phenotypes being often characterized by:
- tissue eosinophilia
- Th2 cytokines
High levels of TSLP have been observed in bronchial epithelial cells derived from asthmatic patients, where the levels of TSLP have been found to be inversely related to lung function.
In a study published in 2011 by the journal Clinical & Experimental Immunology, the authors, with Fang Zhang as lead author, were able to prevent airway inflammation by:
- blocking TSLP signaling through the soluble TSLP R-Ig in a murine asthma model
- down-regulating co-stimulatory molecule expression (CD40, CD80, and CD86) on pulmonary dendritic cells
Moreover, in a study published in 2010 by The Journal of Allergy and Clinical Immunology (JACI), the authors, with Abdelhabib Semlali as lead author, demonstrated that TSLP is able to promote epithelial airway proliferation and epithelial injury repair trough IL13 production.
Epithelial cells from asthmatic patients increased TSLP messenger RNA (ribonucleic acid) or mRNA levels after IL13 stimulation and such cells, after TSLP stimulation, were able to increase IL13 production in a dose-dependent manner.
These findings suggest a positive enhancing loop between TSLP and IL13 in asthmatic patients.
The above data underline that TSLP plays an important role in promoting aberrant Th2 response to antigens.
TSLP And Allergic Rhinitis
The prevalence of allergic rhinitis increased remarkably in the last years with high social costs.
In allergic rhinitis patients, nasal mucosa, after exposure to allergens, is involved by:
- a chronic inflammatory process with increased serum immunoglobulin E (IgE)
- tissue infiltration of eosinophils
- Th2 cells and mucus hyperproduction
It results in a Th1 and Th2 cell imbalance, with hyperproduction of IL4, IL5, and IL13.
In a study published in 2010 by the journal Allergy, the authors, with G. Xu as lead author, demonstrated that TSLP and IL25 were up-regulated in the nasal lavages and in nasal epithelial cells stimulated with polyinosinic:polycytidylic acid (poly I:C).
Moreover, they showed a positive correlation between epithelial cells production of TSLP and IL25, confirming that the epithelial layer is not only a mechanical barrier but a key factor in the initiation and maintenance of the inflammatory environment, characterized by eosinophil recruitment promoted by IL25 and Th2 cytokines.
In a study published in 2009 by the journal Acta Oto-Laryngologica, the authors, with Zhonglin Mou as lead author, demonstrated that TSLP is over expressed in the epithelial layer of nasal mucosa based on:
- immunostaining
- mRNA folds
- concentration protein
These authors also showed a positive correlation between TSLP and IL4 level, strictly linked with allergic rhinitis severity.
The positive correlations observed between TSLP and Th2 cytokines (IL4) and between TSLP and epithelial derived cytokines (IL25) emphasizes the central role of TSLP in allergic rhinitis pathogenesis.
TSLP And Chronic Rhinosinusitis
Chronic rhinosinusitis is a common disease that can significantly decrease quality of lifeand require significant direct medical expense.
The relative roles of initiating events, environmental factors and host susceptibility factors are currently unknown.
Chronic rhinosinusitis is a multifactorial disease, where contributing factors may be:
- mucociliary impairment
- bacterial infection
- allergy
- swelling of the mucosa
- rarely
- physical obstruction caused by anatomical variations in the nasal cavities
- paranasal sinuses
Chronic rhinosinusitis is commonly classified into two phenotypes:
- chronic rhinosinusitis with nasal polyps (CRSwNP)
- chronic rhinosinusitis without nasal polyps (CRSsNP)
Nasal polyps consist of connective tissue, edema, and inflammatory cells like:
- eosinophils (the most common)
- neutrophils
- mast cells
- plasma cells
- lymphocytes
- monocytes
Nasal polyps (CRSwNP) have been associated with high levels of:
a. IL5 (interleukin 5)
b. local nasal immunoglobulin E (IgE) production
c. sinus inflammatory infiltrate rich in:
- eosinophils
- eotaxin
- eosinophil cationic protein (Th2 driven disease)
CRSsNP shows a less defined T helper 2 (Th2) profile and sinus inflammatory infiltrate rich in neutrophils, as well INFγ (interferon-gamma) and transforming growth factor-β (TGFβ) production have been reported.
In a study published in 2011 by the journal The American Journal of the Medical Sciences, the authors, with Tao Liu as lead author, investigated the role of TSLP in nasal polyp development.
These authors demonstrated that thymic stromal lymphopoietin (TSLP) levels in the nasal epithelial layer were higher in patients with nasal polyps compared to patients without nasal polyps, and their data could not be explained by atopy.
In the same study, the authors observed that dendritic cells, obtained from nasal polyp mucosa stimulated with nasal polyp proteins extract, showed higher expression of TSLP R and were able to up-regulate OX40L compared to dendritic cells derived from controls.
These data indicate that the expression of thymic stromal lymphopoietin receptor (TSLP R) makes many cells, apart from dendritic cells, responsive to TSLP produced by nasal epithelial cells, promoting a Th2-inflammatory response.
In a study published in 2011 by the journal Allergy, Asthma & Immunology Research, the authors, with Satoko Kimura as lead author, demonstrated that, compared to non-allergic subjects, immunoreactivity for TSLP was higher in:
- nasal mucosa of allergic rhinitis
- nasal polyps from allergic patients
Moreover, eosinophils count and IgE levels in nasal polyps positively correlate with TSLP expression in several cell types, such as:
- epithelial
- endothelial
- fibroblast
- inflammatory cells
In a study published in 2011 by the International Journal of Immunopathology and Pharmacology, the authors, with Monica Boita as lead author, demonstrated a statistically significant difference in TSLP R expression in CRSwNP and CRSsNP compared to normal subjects, both in the inflammatory and epithelial cells, with no significant difference between chronic rhinosinusitis subtypes (CRSwNP and CRSsNP).
The cells with the highest TSLP R immunostaining score were:
- eosinophils
- mast cells
- histiocytes
The results agree with the data found in a study published in 2007 by the Journal of Experimental Medicine (JEM), with Zoulfia Allakhverdi as lead author.
Allakhverdi et al. showed that eosinophils and mast cells express the functional TSLP R complex and are able to respond to TSLP signals in the presence of an inflammatory environment.
Moreover, we observed that the number of OX40L positive cells, as well as mRNA expression of OX40L, are increased in the lamina propria of sinus tissue of patients with CRSwNP, with a significant correlation with the eosinophilic infiltration compared to CRSsNP (submitted data).
This observation suggests an early common pathway which starts from the epithelial barrier through TSLP-TSLP R as a possible driving factor in the pathogenesis of chronic rhinosinusitis.
The axis TSLP-OX40 should play a major role in promoting Th2 polarization and eosinophilic inflammation only in CRSwNP (submitted data).
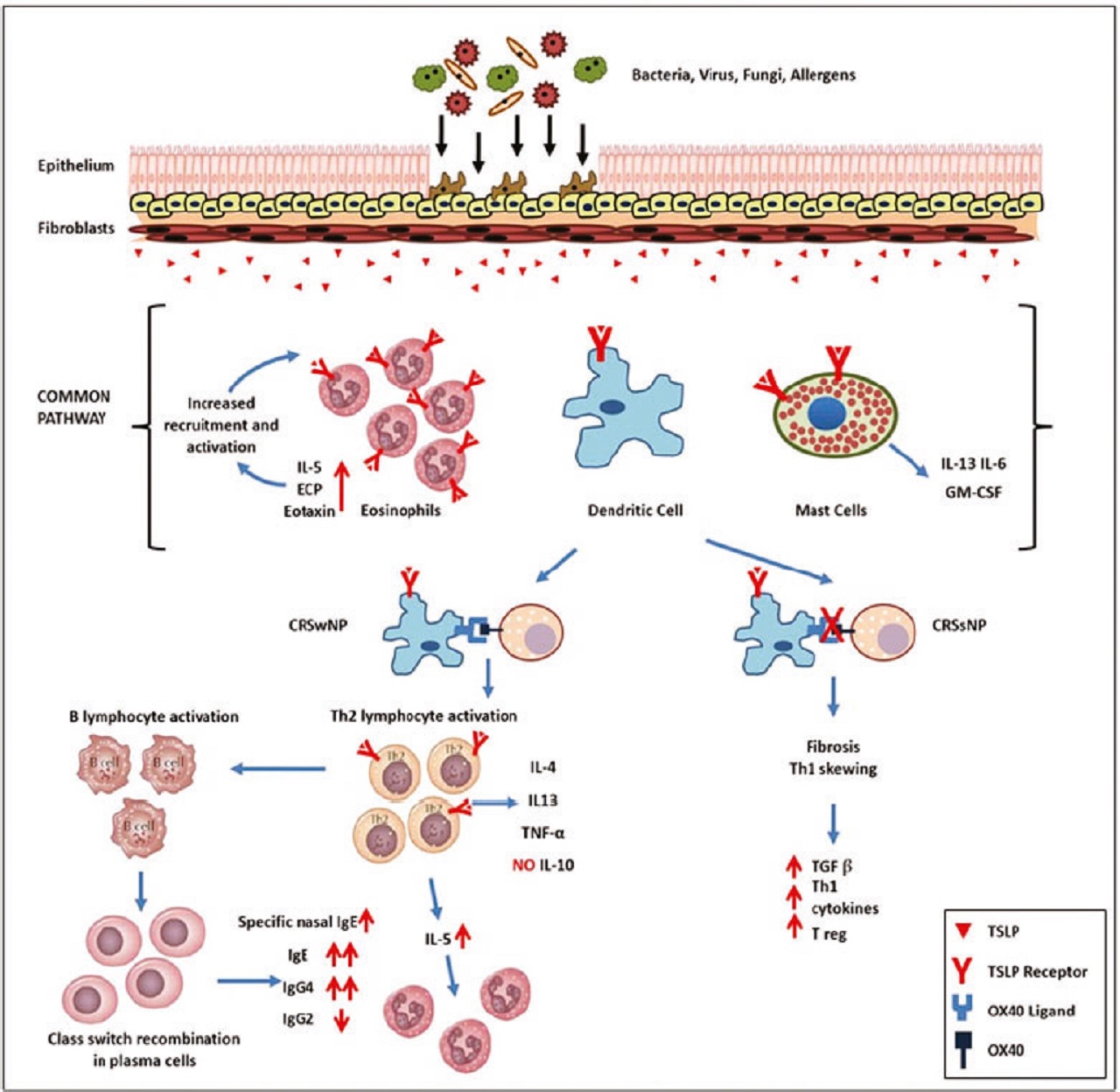
Many data suggest that TSLP plays a central role in nasal polyp development.
Increased TSLP levels have been reported in nasal polyp and TSLP drive dendritic cells towards a Th2 phenotype by upregulating OX40L on dendritic cells surface.
Moreover, TSLP levels positively correlate with eosinophil tissue infiltrate and IgE levels in allergic subjects with nasal polyp.
These findings suggest that TSLP-TSLP R interaction at the level of epithelial layer may play an important role in the pathogenesis of chronic rhinosinusitis characterized by Th2 inflammatory changes of sinus mucosa.
The emerging concept of tissue-specific control of immunity, with TSLP secretion by epithelial cells acting as an initial factor in the chronic rhinosinusitis cascade, can partially explain the early stages of the disease, while the mechanisms involved in the nasal polyp formation are still unknown.
Conclusion
Asthma is a common comorbidity in subjects with allergic rhinitis and chronic rhinosinusitis, and epidemiological surveys have suggested a close connection between upper and lower airway diseases expressed as the “united airways concept.”
The recent findings of thymic stromal lymphopoietin (TSLP) involvement in both upper and lower airway diseases suggest that this cytokine may partially explain the similar T helper 2 (Th2) inflammatory pattern.
According to the 2011 study by Zhang et al., where TSLP signaling was successfully blocked by a soluble TSLP R-Ig in a murine asthma model, TSLP could be considered in the near future as a therapeutic target for biological therapies of asthma and rhinosinusitis.
It recommended that studies relating to thymic stromal lymphopoietin in respiratory disorders should be further conducted.
Jump to
Discussion
TSLP Cytokine
Thymic Stromal Lymphopoietin Receptor
TSLP And Immune Response
Dendritic Cells
Dendritic Cells And OX40/OX40L
TSLP And Adaptive Immunity
TSLP And Innate Immunity
TSLP And Epithelial Cell-Derived Cytokines Network
TSLP And Diseases
TSLP And Atopic Dermatitis
TSLP And Asthma
TSLP And Allergic Rhinitis
TSLP And Chronic Rhinosinusitis
Conclusion

Suleman Shah
Author
Suleman Shah is a researcher and freelance writer. As a researcher, he has worked with MNS University of Agriculture, Multan (Pakistan) and Texas A & M University (USA). He regularly writes science articles and blogs for science news website immersse.com and open access publishers OA Publishing London and Scientific Times. He loves to keep himself updated on scientific developments and convert these developments into everyday language to update the readers about the developments in the scientific era. His primary research focus is Plant sciences, and he contributed to this field by publishing his research in scientific journals and presenting his work at many Conferences.
Shah graduated from the University of Agriculture Faisalabad (Pakistan) and started his professional carrier with Jaffer Agro Services and later with the Agriculture Department of the Government of Pakistan. His research interest compelled and attracted him to proceed with his carrier in Plant sciences research. So, he started his Ph.D. in Soil Science at MNS University of Agriculture Multan (Pakistan). Later, he started working as a visiting scholar with Texas A&M University (USA).
Shah’s experience with big Open Excess publishers like Springers, Frontiers, MDPI, etc., testified to his belief in Open Access as a barrier-removing mechanism between researchers and the readers of their research. Shah believes that Open Access is revolutionizing the publication process and benefitting research in all fields.

Han Ju
Reviewer
Hello! I'm Han Ju, the heart behind World Wide Journals. My life is a unique tapestry woven from the threads of news, spirituality, and science, enriched by melodies from my guitar. Raised amidst tales of the ancient and the arcane, I developed a keen eye for the stories that truly matter. Through my work, I seek to bridge the seen with the unseen, marrying the rigor of science with the depth of spirituality.
Each article at World Wide Journals is a piece of this ongoing quest, blending analysis with personal reflection. Whether exploring quantum frontiers or strumming chords under the stars, my aim is to inspire and provoke thought, inviting you into a world where every discovery is a note in the grand symphony of existence.
Welcome aboard this journey of insight and exploration, where curiosity leads and music guides.
Latest Articles
Popular Articles